3B Scientific Cell Division II Chart, Meiosis – страница 2
Инструкция к 3B Scientific Cell Division II Chart, Meiosis

®
Français Méiose
(séquences courtes, riches en AT de l’ADN) (2), sont visibles. Les chromatides non sœurs dans lesquelles un
échange de segments de gènes a eu lieu sont encore toujours reliées entre elles par des ponts de chiasma (3).
La métaphase I succède à la prophase I. Les phases méiotiques restantes nécessitent moins de 10 % du
temps total d’une méiose complète.
6. Métaphase I
Lors de la transition de la prophase I à la métaphase I, les paires de centrioles (1) sont arrivées aux deux
pôles opposés de la cellule. Un appareil fusorial s'est formé et la membrane nucléaire (2) se dégrade. Les
chromosomes se disposent sur le plan équatorial. Ils forment la plaque de la métaphase. Vus de haut, les
chromosomes apparaissent sous forme d'étoile (monaster ou sphère-mère). Les cinétochores (3) sont des
complexes protéiques déjà formés sur les centromères. Une particularité de la métaphase I méiotique
réside dans le fait que les cinétochores de chaque paire de chromatides-sœurs semblent avoir fusionné.
Les microtubules (4) du fuseau central qui se sont fixés exactement sur les cinétochores de chaque paire
de chromatides-sœurs (5), indiquent par conséquent tous la même direction. Les ponts de chiasma (6)
sont encore toujours préservés. Ils jouent un rôle important pour la disposition correcte des chromosomes
homologues sur le plan équatorial.
Le réticulum endoplasmique (7) et l’appareil de Golgi (8) sont pratiquement décomposés complètement.
7. Anaphase I
Au cours de l’anaphase I de la méiose, les chromosomes homologues (1) se séparent et non les chroma-
tides-sœurs comme au cours de la mitose. Les ponts de chiasma qui retenaient les chromosomes homolo-
gues maternels et paternels se décomposent.
Quelques organismes mutants, chez lesquels l’enjambement méiotique n’est effectué que dans une certaine
mesure, présentent des paires de chromosomes exemptes de ponts de chiasma. La plupart du temps, ces
paires ne sont pas vraiment désunies (non-disjonction), et les cellules filles en résultant possèdent un chro-
mosome trop peu ou un chromosome de trop. On désigne une telle déformation par le terme d’aberration
chromosomique numérique, pouvant entraîner des anomalies.
La séparation commence au niveau des cinétochores (2), lieu de fixation des fibres tractrices du fuseau cen-
tral. C’est à partir de cet endroit que les chromosomes sont lentement tirés par les microtubules (3) se rac-
courcissant, et par l’effet de traction en résultant vers les centrioles (4) se trouvant aux pôles de la cellule.
Les microtubules (5) qui ne sont pas liés à des chromosomes s’allongent, ce par quoi la distance entre les
centrioles augmente, et la cellule s’étire dans le sens de la longueur. Sur le plan équatorial, on reconnaît
l’ébauche d’un sillon d’étranglement (6).
Par l’enjambement effectué au cours de la prophase et la répartition fortuite des chromosomes maternels
et paternels vers les deux pôles de la cellule, on observe une variation du patrimoine héréditaire (voir
introduction).
8. Télophase I, Cytocinèse I, Intercinèse, Prophase II et Métaphase II
Télophase I et Cytocinèse
Au cours de la télophase I, l’appareil fusorial se résorbe, et un étranglement annulaire (1) se forme au
niveau du plan équatorial. De même, une fine membrane cellulaire (2) se forme. Au cours de la cytocinèse
suivante, le corps cellulaire est divisé en deux parties exactes au centre, c'est-à-dire à l'étranglement annu-
laire entre les deux nouveaux noyaux fils (3). Chaque noyau fils contient le jeu de chromosomes maternel,
resp. paternel légèrement modifié par l’enjambement, l’ADN est déjà dédoublé, c.-à-d. qu’un chromosome
se compose de deux chromatides-sœurs (4).
21

®
FrançaisMéiose
Le réticulum endoplasmique (5) et l’appareil de Golgi (6) ont retrouvé leur forme et leur taille initiales.
A la fin de la cytocinèse, la première division de maturation est achevée.
Intercinèse
La première et la deuxième division de maturation sont séparées par une courte phase de repos (interphase).
Un dédoublement des chromosomes issus de deux chromatides n’a cependant pas lieu (pas de phase S).
Les deux chromatides-sœurs de chaque chromosome restent liées entre elles par les centromères.
Division de maturation II
La deuxième division de maturation se déroule comme une mitose (division habituelle du noyau et de la
cellule). C’est la raison pour laquelle on l’appelle également division équationnelle. Etant donné que les
chromosomes n’ont pas été redédoublés au cours de l'intercinèse précédente, on observe une réduction
du patrimoine héréditaire du jeu de chromosomes haploïde lors de la seconde division méiotique y succédant.
Prophase II
La prophase II correspond étroitement à la prophase d’une mitose et se déroule très rapidement dans
tous les organismes. La perméabilité de la surface de la cellule augmente afin d’absorber le liquide de
l’environnement. L’appareil microtubulaire du cytosquelette se restructure. La membrane nucléaire se
désagrège, et le fuseau se forme à partir de microtubules se restructurant.
Métaphase II
Au cours de la métaphase II, les chromosomes sont à nouveau disposés sur le plan équatorial, et les deux
extrémités du fuseau se trouvent aux deux pôles opposés l’un à l’autre (de façon analogue à la métaphase I).
Une différence essentielle par rapport à la métaphase I réside dans le fait que deux cinétochores se sont
développés sur les chromatides-sœurs qui cette fois indiquent des directions polaires contraires.
9. Anaphase II
Au cours de l’anaphase II, les deux chromatides-sœurs (1) de chaque chromosome sont séparées comme au
cours de la mitose.
La séparation commence au niveau des cinétochores (2), au lieu de fixation des fibres tractrices du fuseau
central. A partir de cet endroit, les chromatides sont tirées lentement par les microtubules (3) se raccourcis-
sant, et par l’effet de traction en résultant vers les centrioles (4) se trouvant aux pôles de la cellule.
Les microtubules (5) qui ne sont pas liés à des chromatides s’allongent, ce par quoi la distance entre les
centrioles augmente, et la cellule s'étire dans le sens de la longueur. Sur le plan équatorial, on reconnaît
l’ébauche d’un sillon d’étranglement (6).
10. Télophase II et Cytocinèse II
De l’étranglement et de la séparation des deux cellules issues de la première division de maturation
résultent maintenant quatre cellules haploïdes (1) possédant une combinaison de gènes différente, causée
par la répartition fortuite des chromosomes et par l’enjambement. Ceci permet d’expliquer le fait que
frères et sœurs ne sont pas identiques : un enfant présente plus les caractéristiques du père, l’autre enfant
présente plus les caractéristiques de la mère. C’est pourquoi des caractéristiques appartenant aux ascen-
dants peuvent réapparaître.
22
1
2
6
1
7
10
3
4
8
5
9
2
3
2
4
5
1
4
3
4
1
2
3
23
4
1
1
2
5
2
3
1
6
1
8
5
6
3
4
2
7
1
24
7
4
6
1
5
2
3
4
8
2
7
3
1
6
4
5
1
9
2
4
6
5
3
25
10
1
1
1
1
26

®
Português
Meiose
A meiose, também chamada de divisão reducional, é uma forma especial de divisão celular. Dependendo
do organismo ou do tipo de célula, ela pode durar poucos dias ou também anos. O processo de meiose
origina gametas (células germinativas), ou seja, a divisão reducional de uma célula precursora leva à
formação de quatro gametas. Basicamente, podemos diferenciar dois tipos de gametas. Gametas grandes
e imóveis são chamados de óvulos ou oócitos e gametas pequenos e móveis são chamados de espermato-
zóides.
Os óvulos são formados nos ovários a partir de células precursoras por meio da divisão meiótica. A matu-
ração dos óvulos do ser humano inicia-se já no estado embrionário (3o a 4o mês de gestação), porém, as
células permanecem em um estado meiótico intermediário até a maturação sexual do organismo. A partir
deste momento, alguns dos óvulos imaturos concluem o processo de divisão meiótica em períodos regu-
lares, controlados por hormônios. Após o início da puberdade, a maturação dos espermatozóides humanos
nos testículos ocorre em intervalos regulares. Neste caso o processo completo da divisão meiótica dura
entre 20 e 24 dias.
Normalmente as células do corpo humano (p. ex. as células precursoras dos gametas) contêm um número
diplóide de cromossomos, ou seja, os cromossomos ocorrem aos pares, sendo um componente do par de
origem paterna e o outro de origem materna. Portanto, cada cromossomo possui uma cópia, sendo parte
de um par de cromossomos correspondentes (homólogos). Os gametas, em contrapartida, são haplóides.
Isto significa que contêm somente um representante de cada par. Portanto, os óvulos ou espermatozóides
possuem apenas a metade dos genes maternos ou paternos; desta maneira, a fusão de ambas as células
leva à formação de uma nova célula (zigoto) com um número de cromossomos diplóide.
O objetivo da meiose consiste em reduzir o número diplóide de cromossomos de uma célula precursora
dos gametas a um número haplóide, a fim de criar as condições prévias para a reprodução sexual. Uma
outra função importante da meiose é a mistura dos genes, obtida através de dois mecanismos:
1. a distribuição aleatória dos cromossomos maternos e paternos às células germinativas formadas;
2. a troca de genes entre os cromossomos homólogos (recombinação genética).
(Os respectivos processos destes dois mecanismos serão expostos mais abaixo no âmbito da descrição das
fases individuais.)
No caso do ser humano, que possui um número haplóide de 23 cromossomos, a distribuição aleatória dos
23
6
cromossomos resulta em um número de variações genéticas possíveis de 2
ou 8,4 x 10
. Esta variabilida-
de genética é adicionalmente aumentada devido à troca de genes entre os cromossomos.
Antes da divisão meiótica, as células precursoras dos gametas encontram-se no estado da interfase, que
corresponde ao período intermediário entre duas divisões celulares (mitóticas ou meióticas). A interfase
abrange 3 etapas:
• Fase G
(pré-síntese) Esta fase é caracterizada pelo crescimento da célula.
1
• Fase S (síntese) Nesta etapa inicia-se a duplicação dos centríolos e do DNA (ácido desoxirribonucléico).
• Fase G
(pós-síntese) Esta fase marca o fim da síntese de DNA antes da fase de divisão. Além disto, é
2
concluído o processo de duplicação dos centríolos.
A interfase é seguida pela meiose, que consta de duas fases de divisão separadas por uma curta interfase
especial (intercinese). Como também é o caso na mitose, cada uma destas divisões é subdividida em dife-
rentes etapas:
27

®
PortuguêsMeiose
Divisão I
• Prófase I (abrange quatro subetapas: leptóteno, zigóteno, paquíteno e diplóteno com diacinese)
• Metáfase I
• Anáfase I
• Telófase I
• Citocinese I
Intercinese
Divisão II
• Prófase II
• Metáfase II
• Anáfase II
• Telófase II
• Citocinese II
A série de modelos sobre a meiose da 3B Scientific® (número de produto: R02) e o quadro esquemático da
mitose (V2051M, V2051U) mostra uma célula animal típica em uma escala de aproximadamente 10.000:1.
Na parte inferior dos esquemas encontram-se cortes das organelas celulares.
A série de modelos sobre a meiose da 3B Scientific® é fornecida com uma embalagem equipada com um
dispositivo de suspensão, com o qual é possível pendurar a série de modelos na parede para economizar
espaço. Ímãs fixados no reverso dos modelos possibilitam a sua disposição em quadros magnéticos na sala
de aula.
Ao final desta apresentação, você encontrará esquemas das 10 etapas apresentadas, que podem ser copi-
ados e utilizados em aula. Pintando os esquemas, preenchendo as legendas e colocando as diferentes eta-
pas na ordem certa, seus alunos poderão facilmente entender e aprofundar o que aprenderam.
Esquemas coloridos das diferentes etapas podem ser também gratuitamente obtidos na Internet sob o
endereço www.3bscientific.com.
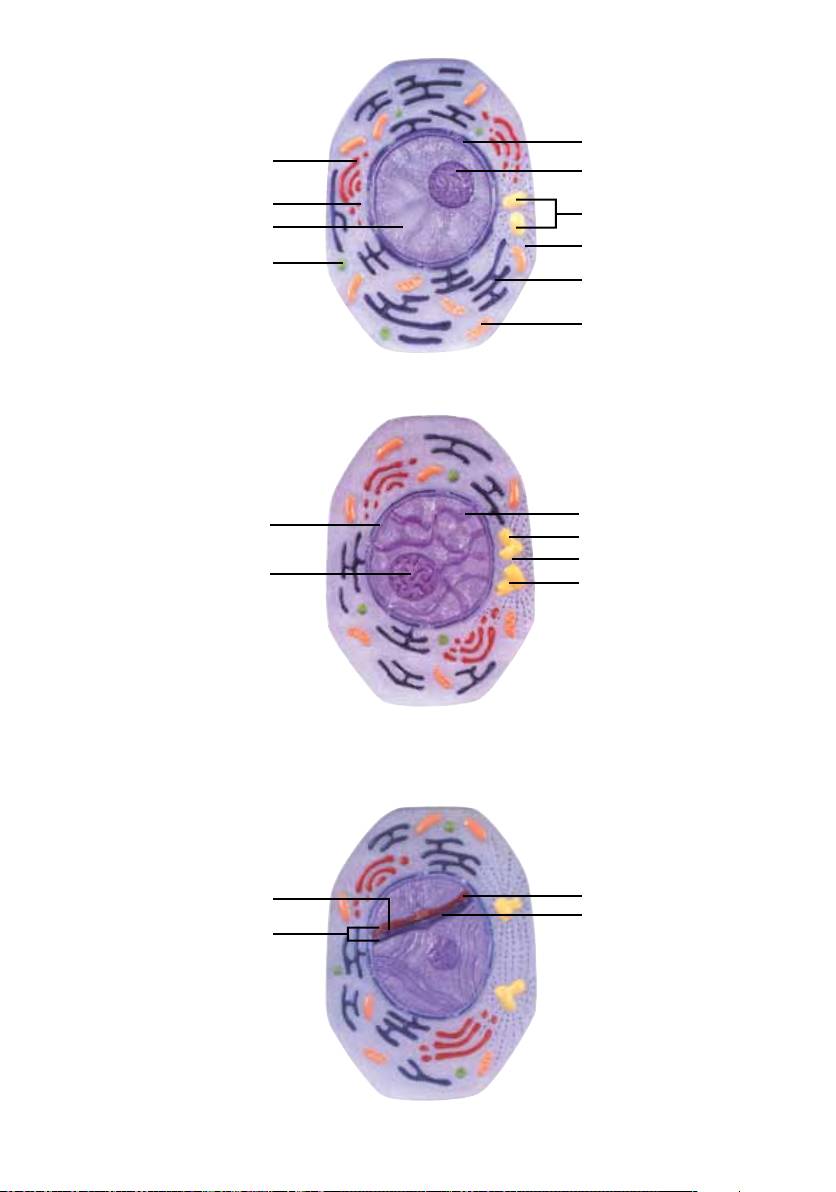
1. Interfase, fase G
1
No interior da célula, podemos observar o núcleo celular com o nucléolo (1) e a carioteca (membrana
nuclear) (2). Dentro do núcleo celular encontra-se o DNA ainda não condensado (3) com a informação
genética.
A estabilidade e a forma da célula são garantidos através de cilindros extremamente finos, os chamados
microtúbulos (4) presentes no citoplasma. Os microtúbulos também são responsáveis pelo movimento da
célula e pelos processos de transporte intracelulares.
Dentro do citoplasma podemos observar o retículo endoplasmático (5). Trata-se de uma complexa rede
de tubos, responsável sobretudo pela síntese de lipídios e pelo transporte de íons e certas proteínas no
interior da célula. Junto à membrana do retículo endoplasmático encontram-se os ribossomos, cuja função
consiste na produção de proteínas.
O complexo de Golgi (6) é também chamado de “glândula celular”. Ele é constituído por empilhamentos de
pequenos sacos achatados, em cujas bordas brotam pequenas vesículas (vesículas de Golgi) (7). O complexo
de Golgi recebe enzimas e componentes do retículo endoplasmático para a formação de suas membranas.
Sua função principal consiste no armazenamento e transporte de secreções e na síntese de lisossomos
(vesículas cheias de enzimas digestivas) (8).
A função principal dos lisossomos é a digestão de material celular. Esta digestão pode ocorrer de dentro
para fora da célula (=exocitose) ou dentro da própria célula (=digestão intracelular, endocitose). A energia
dentro da célula é gerada pelas mitocôndrias (9).
O papel dos centríolos (10) consiste em formar o fuso de divisão. Os centríolos têm a forma de cilindros
ocos, formados por túbulos longitudinais (microtúbulos).
28

®
Português
Meiose
2. Prófase I
A prófase da primeira divisão é o período de maior duração da meiose. Durante esta etapa, os cromosso-
mos e a cromatina alteram a sua estrutura e sua posição no núcleo conforme uma determinada seqüência.
Por esta razão, a prófase I é subdividida em quatro períodos (leptóteno, zigóteno, paquíteno, diplóteno
com diacinese). Ao contrário da prófase mitótica, que tem uma duração de poucas horas, a prófase I da
meiose pode demorar vários dias, semanas, meses ou anos.
Leptóteno
No início da prófase I (leptóteno), podemos observar o nucléolo (1) e a carioteca (membrana nuclear) (2).
Os cromossomos (3) tornam-se visíveis como filamentos finos, compridos e individualizados, cujas extre-
midades estão fixadas à carioteca. Os cromossomos replicados (duplicados) durante a interfase precedente
consistem em duas cromátides irmãs que encontram-se muito próximas uma à outra, tornando impossível
a sua distinção.
Os centríolos também são duplicados durante a interfase. Ambos os pares (4) começam a afastar-se grada-
tivamente em direção aos pólos da célula. Entre eles forma-se o chamado fuso meiótico (5), composto por
numerosos microtúbulos.
3. Zigóteno e Paquíteno
O modelo mostra respectivamente um cromossomo homólogo materno (1) e paterno (2) (compostos por
duas cromátides irmãs) de um par de cromossomos, representados em diferentes cores, simbolizando o
conjunto completo de cromossomos (ao todo 2 x 23).
Zigóteno
A etapa do zigóteno inicia-se a partir do momento em que os cromossomos homólogos começarem a se
juntar, formando o complexo sinaptonemático (3) (disposição paralela dos pares homólogos). Em geral,
este processo começa em uma extremidade dos cromossomos e é continuado progressivamente até chegar
à outra extremidade, semelhante a um zíper. Este pareamento dos cromossomos (sinapse) ocorre com
grande precisão, sendo que ao final do processo os genes correspondentes dos cromossomos homólogos
encontram-se diretamente opostos. Este é um pré-requisito para a recombinação de pedaços de genes
(crossing over). Na prófase I da meiose, cada par de cromossomos homólogos é chamado de bivalente;
porém, pelo fato de que cada cromossomo homólogo individual é constituído por duas cromátides irmãs,
o conjunto destes quatro filamentos de cromossomos homólogos pareados é também chamado de tétrade.
Paquíteno
Após a formação completa de todos os complexos synaptonemáticos, isto é, após a conclusão do parea-
mento dos cromossomos homólogos, inicia-se a fase do paquíteno. Nos complexos synaptonemáticos tor-
nam-se visíveis os pontos de recombinação (4), nos quais será realizada a troca de segmentos de genes.
4. Diplóteno
Após as quebras em cromátides homólogas seguidas de soldadura de pedaços de genes trocados, os cro-
mossomos homólogos (1) começam a afastar-se gradativamente, permanecendo em contato em alguns
pontos chamados quiasmas (2). Os quiasmas representam as regiões em que houve a recombinação genéti-
ca (troca de informação genética materna e paterna). Os óvulos podem permanecer no estado do diplóteno
durante vários meses ou anos.
5. Diacinese
O final da prófase I da meiose inicia-se com a separação dos cromossomos da carioteca (1). A condensação
dos cromossomos aumenta ainda mais e as cromátides irmãs, ligadas através dos centrômeros (seqüências
de DNA curtas e ricas em AT) (2), tornam-se visíveis. As cromátides não irmãs, ou seja, aquelas em que
houve uma troca de segmentos de genes, permanecem em contato nos quiasmas (3).
29

®
PortuguêsMeiose
O final da prófase I marca o início da metáfase I. As fases meióticas restantes têm uma duração de menos
de 10% da duração total de uma meiose completa.
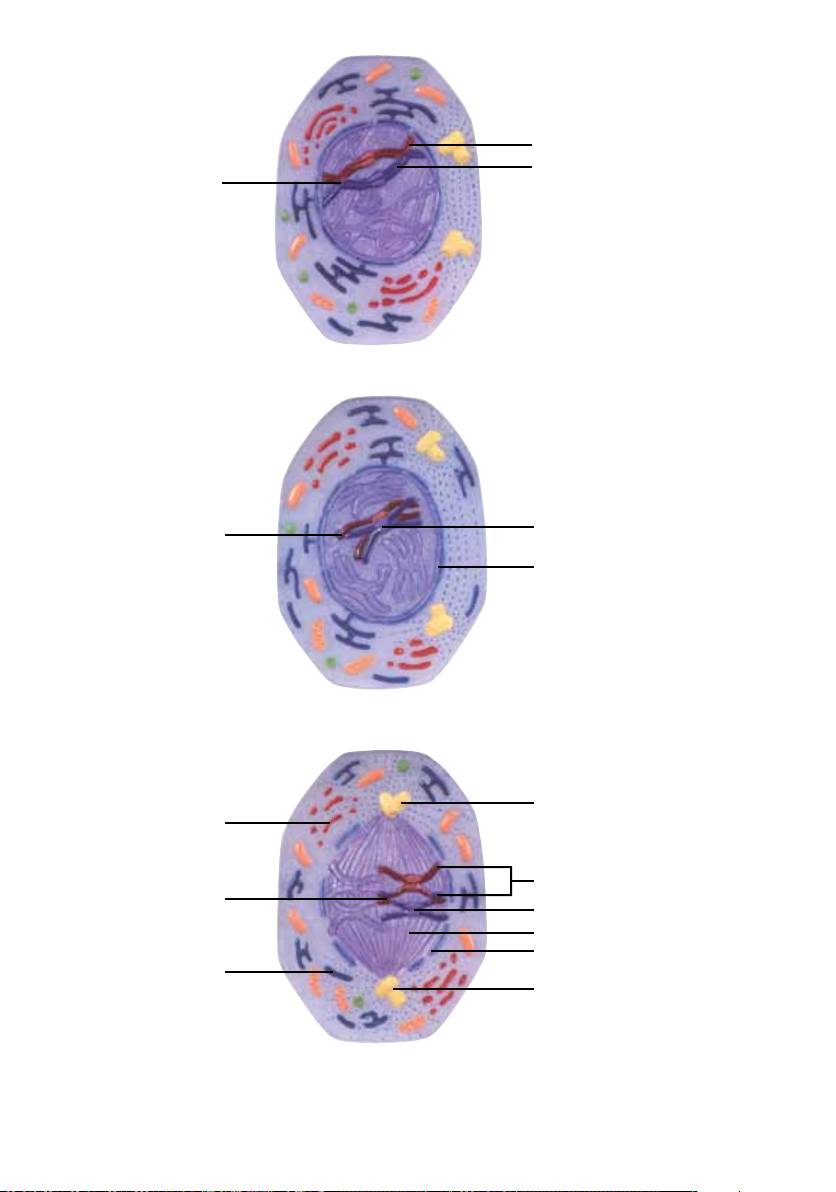
6. Metáfase I
No final da prófase I e no início da metáfase I, os pares de centríolos (1) chegaram aos dois pólos opostos
da célula. Formou-se um fuso e a carioteca (2) se dissolve. Os cromossomos se organizam no plano equatorial
da célula. Eles formam a chamada placa metafásica (placa equatorial). Vistos de cima, os cromossomos
apresentam um aspecto de estrela (monáster ou estrela-mãe). Os cinetócoros (3) são complexos de proteína
que se formaram nos centrômeros. Um aspecto específico da metáfase I da meiose é o fato de os cinetócoros
de cada par de cromátides irmãs parecerem estar fusionados. Desta maneira, os microtúbulos (4) do fuso
meiótico, que se ligaram exatamente aos cinetócoros de cada par de cromátides irmãs (5), apontam todos
para a mesma direção. Os quiasmas (6) ainda permanecem existentes. Eles desempenham um papel central
para a organização correta dos cromossomos homólogos no plano equatorial.
O retículo endoplasmático (7) e o complexo de Golgi (8) estão quase completamente desintegrados.
7. Anáfase I
Durante a anáfase I da meiose ocorre a separação dos cromossomos homólogos (1) e não, como é o caso
na mitose, das cromátides irmãs. Os quiasmas, que até este momento haviam ligado os cromossomos
homólogos maternos e paternos, desaparecem.
Alguns organismos mutantes, em que o crossing over meiótico ocorre somente em medida limitada,
apresentam pares de cromossomos sem quiasmas. Em geral, estes pares não são separados corretamente
(nondisjunction) e as células-filhas resultantes contêm um cromossomo a mais ou a menos. Uma mutação
deste tipo é chamada de aberração cromossômica numérica, que resulta em malformações.
A separação inicia-se nos cinetócoros (2), os pontos aos quais se prendem as fibras do fuso de divisão.
Através do encurtamento dos microtúbulos (3) e o efeito de tração resultante, os cromossomos migram
em direção aos centríolos (4) localizados nos pólos da célula. Os microtúbulos (5) que não estão ligados a
cromossomos tornam-se mais longos, aumentando a distância entre os centríolos e tornando a célula mais
longa. No plano equatorial da célula começa a aparecer uma constrição (6).
Devido ao crossing over na pró-fase e a distribuição aleatória dos cromossomos maternos e paternos aos
pólos da célula é atingida uma grande variabilidade genética (veja introdução).
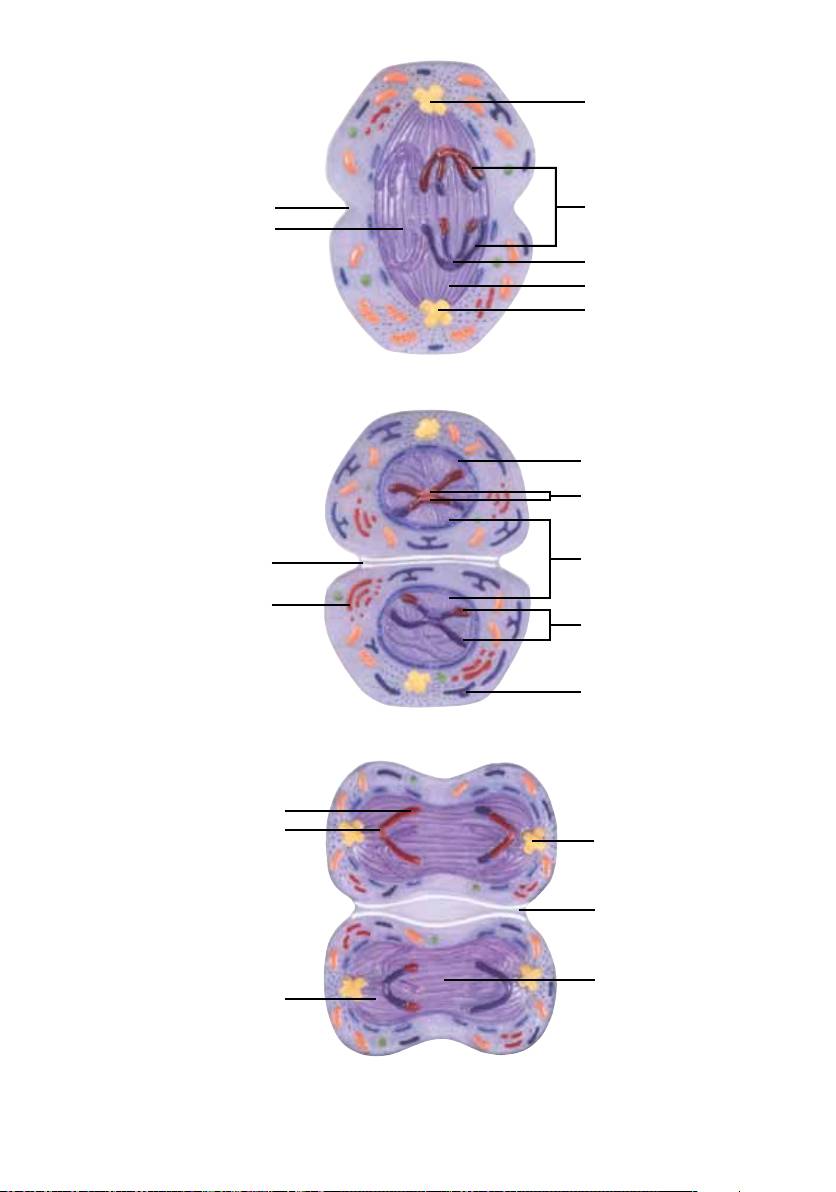
8. Telófase I, Citocinese I, Intercinese, Prófase II e Metáfase II
Telófase I e Citocinese
Durante a telófase I, o fuso começa a desaparecer e uma constrição (1) forma-se no plano equatorial. Além
disto, é formada uma carioteca fina (2). Durante a citocinese que segue, o corpo celular é dividido exatamente
ao meio, na constrição entre os dois núcleos filhos recém-formados (3). Através do crossing over, os
núcleos-filhos recebem cromossomos maternos e paternos diferentes, sendo que o DNA já foi duplicado,
isto é, um cromossomo é composto por duas cromátides irmãs (4).
O retículo endoplasmático (5) e o complexo de Golgi (6) voltam a adquirir a sua forma e seu tamanho original.
O final da citocinese corresponde ao fim da primeira divisão.
Intercinese
A primeira e a segunda divisão são separadas por uma curta fase de repouso (interfase). Porém, não ocorre
uma duplicação dos cromossomos compostos por duas cromátides (não há uma fase S). As cromátides
30

®
Português Meiose
irmãs de cada cromossomo permanecem ligadas através dos centrômeros (7).
Divisão II
A segunda etapa de divisão corresponde ao processo da mitose (divisão celular e nuclear normal). Por esta
razão, ela é também chamada de divisão equacional. Sendo que os cromossomos não foram novamente
duplicados durante a intercinese, a segunda divisão meiótica resulta na redução dos genes ao número
haplóide de cromossomos.
Prófase II
A prófase II é uma prófase semelhante à da mitose e é muito rápida em todos os organismos. A permeabilidade
da membrana celular aumenta, possibilitando a absorção de líquidos. O complexo de microtúbulos do
citoesqueleto é reorganizado. A carioteca desaparece e o fuso é novamente formado a partir dos microtú-
bulos reorganizados.
Metáfase II
Durante a metáfase II, os cromossomos se posicionam novamente no plano equatorial da célula e as duas
extremidades do fuso migram para os pólos opostos da célula (igual à metáfase I). Uma diferença impor-
tante em relação à metáfase I consiste no fato de que nas cromátides irmãs se desenvolveram dois cinetó-
coros, que agora estão orientados para pólos diferentes, em direção oposta.
9. Anáfase II
Segue a anáfase II, durante a qual as duas cromátides irmãs (1) de cada cromossomo são separadas, como
é também o caso na mitose.
A separação inicia-se nos cinetócoros (2), aos quais se prendem as fibras do fuso de divisão. Através do
encurtamento dos microtúbulos (3) e o efeito de tração resultante, os cromossomos são arrastados gradual-
mente em direção aos centríolos (4) localizados nos pólos da célula. Os microtúbulos (5) que não estão liga-
dos a cromátides tornam-se mais compridos, aumentando assim a distância entre os centríolos e tornando
a célula mais longa. No plano equatorial, começa a aparecer uma constrição (6).
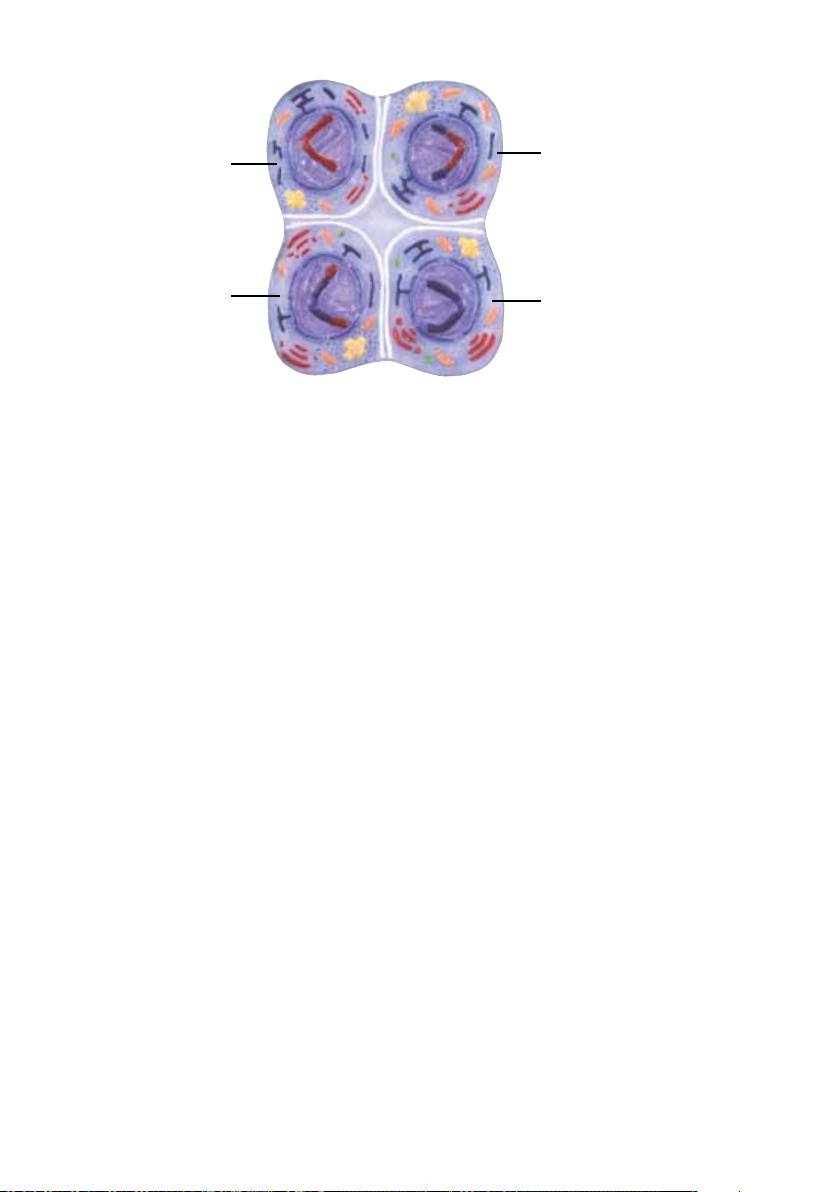
10. Telófase II e citocinese II
Através da constrição e divisão das duas células resultantes da primeira divisão meiótica formam-se quatro
células haplóides (1) com diferentes combinações genéticas causadas pela distribuição aleatória dos
cromossomos e pelo crossing over. Isto explica o fato de irmãos nunca serem idênticos: um filho apresenta
mais características do pai, o outro tem a aparência da mãe. E desta maneira também podem reaparecer
características de outros parentes antepassados.
31

®
Meiosi
EnglishItaliano
La meiosi, o divisione riduzionale, è un tipo particolare di divisione cellulare. A seconda dell’organismo e
della varietà di cellula interessata, può durare da alcuni giorni ad alcuni anni e ha come risultato la pro-
duzione di cellule sessuali (gameti). Ogni cellula precursore produce quattro gameti attraverso la divisione
riduzionale. In generale, esistono due tipi di gameti. Le grandi cellule immobili vengono definite cellule
uovo od ovociti, mentre i gameti piccoli e mobili sono gli spermatozoi o spermatociti.
Le cellule uovo vengono create attraverso la divisione meiotica da parte di cellule precursori nelle ovaie.
Benché le cellule uovo umane inizino a maturare già nell’embrione (dal 3° all’8° mese di gravidanza),
esse restano in una fase meiotica intermedia fino al raggiungimento della maturità sessuale. Da questo
momento in poi, alcune cellule uovo immature completano la divisione meiotica a intervalli regolari sotto
il controllo degli ormoni. La maturazione degli spermatozoi umani avviene regolarmente nei testicoli una
volta raggiunta la maturità sessuale. In questo caso, una divisione meiotica completa richiede 20-24 giorni.
Di solito, le cellule somatiche, cioè i precursori dei gameti, contengono due assetti cromosomici e vengono
perciò definite diploidi: metà dei cromosomi deriva dalla madre e l‘altra metà dal padre. Di conseguenza,
esiste una copia esatta di ciascun cromosoma, ossia, una coppia corrispondente (omologa) di cromosomi.
Al contrario, i gameti contengono un singolo assetto cromosomico e vengono quindi definiti aploidi. In
altre parole, le cellule uovo e gli spermatozoi contengono solo metà del patrimonio genetico della madre e
del padre, così che quando si uniscono, danno origine a una nuova cellula (zigote) che presenta un assetto
cromosomico diploide completo.
Lo scopo della meiosi è ridurre l‘assetto cromosomico normalmente diploide di una cellula precursore
di un gamete trasformandolo in un assetto aploide, per stabilire così la base della riproduzione sessuale.
Un‘altra importante funzione della meiosi è di mischiare le informazioni genetiche. Tale obiettivo viene
raggiunto tramite due meccanismi:
1. una distribuzione casuale dei cromosomi materni e paterni alle cellule sessuali che vengono prodotte
2. lo scambio dei geni tra i cromosomi omologhi (ricombinazione genetica)
Le procedure alla base di questi due meccanismi sono spiegate più avanti nella descrizione delle singole
fasi.
Negli umani, che dispongono di 23 cromosomi nell’assetto aploide, la sola distribuzione casuale dei cro-
mosomi rende possibili 223, ossia 8,4 x 106 diverse varianti genetiche. Il numero di varianti aumenta ulte-
riormente grazie allo scambio di geni tra i cromosomi.
Prima della divisione meiotica, le cellule precursori dei gameti si trovano nell‘interfase, che si riferisce al
periodo tra due divisioni cellulari (mitotica o meiotica). L’interfase comprende tre stadi:
• Fase G
(lacuna presintetica), ossia la fase in cui la cellula cresce.
1
• Fase S (sintesi), in cui i centrioli e il DNA (acido desossiribonucleico) iniziano a duplicarsi.
• Fase G
(lacuna postsintetica), che separa la conclusione della sintesi del DNA dalla fase di divisione.
2
Viene inoltre completata la duplicazione dei centrioli.
La meiosi, stadio che segue l’interfase, comprende due successive divisioni (meiotiche) di maturazione,
separate da una breve interfase specifica (intercinesi). Come nella mitosi, anche nella divisione meiotica si
riconoscono diversi stadi:
32

®
EnglishItaliano Meiosi
Prima divisione meiotica:
• Profase I (quattro sottostadi: leptotene, zigotene, pachitene e diplotene con diacinesi)
• Metafase I
• Anafase I
• Telofase I
• Citocinesi I
Intercinesi
Seconda divisione meiotica:
• Profase II
• Metafase II
• Anafase II
• Telofase II
• Citocinesi II
La serie di modelli 3B Scientific® sulla meiosi (codice prodotto R02) e il tabellone murale sulla meiosi
(V2051M, V2051U) mostrano una tipica cellula mammifera con un fattore di ingrandimento pari a circa
10.000. Nella parte inferiore dei modelli e delle illustrazioni, gli organelli cellulari vengono visualizzati
come se fossero aperti.
La serie viene fornita in un supporto per il deposito, dotato di un dispositivo che permette di appendere i
modelli al muro, così da risparmiare spazio. Questi ultimi sono inoltre dotati di calamite sul retro che per-
mettono di applicarli e disporli su lavagne magnetiche in classe e utilizzarli a fini educativi.
Al termine di questa descrizione seguono le illustrazioni delle 10 fasi, che possono essere fotocopiate per
le lezioni. Colorando, denominando e ordinando le singole fasi nel modo corretto, gli studenti potranno
facilmente ripassare e memorizzare quanto appreso.
Illustrazioni a colori gratuite dei vari stadi sono disponibili anche su Internet all‘indirizzo www.3bscientific.
com.
1. Interfase, stadio della fase G
1
All’interno della cellula è possibile scorgere il nucleo con il nucleolo (1) e la membrana nucleare (2). Il
nucleo contiene inoltre il DNA (3) non ancora elicoidale con le informazioni genetiche.
La cellula riceve la propria stabilità e forma da tubi molto sottili, i cosiddetti microtubuli (4), che si esten-
dono attraverso il citoplasma. Essi controllano, tra le altre cose, i movimenti della cellula e i processi di
trasporto intracellulari.
All’interno del citoplasma è visibile il reticolo endoplasmatico (5). Esso è costituito da un sistema di tubi
intrecciati e destinati alla sintesi dei lipidi, all‘immagazzinamento di ioni, alla trasformazione e al traspor-
to di determinate proteine. La membrana del reticolo endoplasmatico rugoso è ricca di ribosomi, la cui
funzione è sintetizzare le proteine che attraversano il reticolo stesso.
L’apparato del Golgi (6) può essere definito la “ghiandola della cellula”. È costituito da una serie di sacche
cave impilate (cisterne membranose appiattite) che si gonfiano man mano che le vescicole del Golgi (7) si
rimpiccioliscono e si sciolgono. L’apparato del Golgi riceve componenti membranosi ed enzimi dal reticolo
endoplasmatico: la sua funzione principale è di raccogliere e distribuire secrezioni, oltre che di produrre
lisosomi (vescicole digestive) (8).
Lo scopo principale dei lisosomi è disgregare i componenti della cellula (digestione intracellulare).
33

®
ItalianoMeiosi
Gli organelli addetti alla produzione di energia sono i mitocondri (9).
La funzione dei centrioli (10) è assemblare il fuso mitotico. Si tratta di cilindri cavi costituiti da tubi disposti
longitudinalmente (microtubuli).
2. Profase I
La profase della prima divisione meiotica è la parte del processo che richiede più tempo. Nel corso del
suo svolgimento, i cromosomi e la cromatina cambiano la propria struttura e disposizione all‘interno del
nucleo, seguendo un ordine specifico. Di conseguenza, la profase I è suddivisa in quattro sottostadi (lepto-
tene, zigotene, pachitene e diplotene con diacinesi). Al contrario della profase mitotica, che dura diverse
ore, la profase meiotica I può durare giorni, settimane, mesi o anni.
Leptotene
All’inizio della profase I (leptotene), è possibile riconoscere il nucleolo (1) e la membrana nucleare (2).
I cromosomi (3) sono visibili sotto forma di individuali filamenti lunghi e sottili. Le loro estremità sono
collegate all’interno della membrana nucleare. Ogni cromosoma è già stato replicato, ossia duplicato, nel
corso dell‘interfase ed è costituito da due cromatidi fratelli, che sono tuttavia talmente vicino l‘uno all‘altro
che non è possibile differenziarli. Anche i centrioli sono stati duplicati durante l’interfase. Entrambe le
coppie (4) iniziano a muoversi in direzioni opposte verso i due poli della cellula. Tra di essi si forma ciò che
viene definito fuso centrale (5), costituito da numerosi microtubuli.
3. Zigotene e pachitene
Un omologo materno (1) e uno paterno (2), costituiti da due cromatidi fratelli, di una coppia di cromosomi
sono visualizzati con colori diversi per rappresentare gli altri cromosomi (2 x 23 in totale).
Zigotene
Lo zigotene inizia non appena i cromosomi omologhi cominciano ad allinearsi gli uni accanto agli altri per
formare il complesso sinaptinemale (3), con una disposizione parallela a quella dei loro partner omologhi.
Questo processo si verifica solitamente a partire da un‘estremità dei cromosomi e continua fino all’altra
estremità, in modo analogo a una chiusura lampo. L’appaiamento dei cromosomi (sinapsi) avviene in
modo molto preciso, in modo che i geni corrispondenti dei cromosomi omologhi siano rivolti uno diretta-
mente di fronte all‘altro. Questo è un requisito importante per lo scambio ricombinante delle sezioni gene-
tiche (crossing-over). Le coppie di cromosomi omologhi nella profase meiotica I vengono di norma definite
bivalenti, ma poiché ciascun cromosoma omologo è costituito da cromatidi fratelli vicini gli uni agli altri,
possono essere chiamate anche tetradi.
Pachitene
Il pachitene inizia non appena tutti i complessi sinaptinemali sono completamente sviluppati, ossia i cro-
mosomi omologhi sono allineati. Da questo momento in poi, sui complessi sinaptinemali, dove avviene lo
scambio delle sezioni genetiche, i noduli di ricombinazione (4) diventano visibili a intervalli.
4. Diplotene
Dopo lo scambio di alcune sezioni genetiche, i cromosomi omologhi (1) si separano sempre più, restando
collegati in corrispondenza di uno o più punti del crossing-over (chiasmi) (2). I chiasmi cromosomici sono
i punti in cui è avvenuta in precedenza la ricombinazione genetica, ossia lo scambio delle informazioni
genetiche materne e paterne. Le cellule uovo possono restare nello stadio diplotene per mesi, o addirittura
per anni.
5. Diacinesi
La profase meiotica I volge al termine quando i cromosomi si staccano dalla membrana nucleare (1). I
cromosomi sono condensati e diventano visibili i cromatidi fratelli, uniti dal centromero (brevi sequenze di
DNA con un elevato livello di AT)(2). I cromatidi non fratelli in cui è avvenuto uno scambio di sezioni gene-
tiche restano collegati per mezzo dei chiasmi (3).
34

®
Italiano Meiosi
La fase successiva alla profase I è la metafase I. Le fasi meiotiche rimanenti a questo punto richiedono
meno del 10% del tempo totale necessario per una meiosi completa.
6. Metafase I
Durante la transizione dalla profase I alla metafase I, le coppie di centrioli (1) hanno raggiunto i due poli
opposti della cellula. Si è sviluppato un apparato fusolare e la membrana nucleare (2) si dissolve. I cromo-
somi si allineano a livello equatoriale, formando la cosiddetta piastra metafasica. Visti dall’alto assumono
una forma simile a una stella (monaster o stella “madre”). I cinetocori (3) sono complessi di proteine già
sviluppati in corrispondenza dei centromeri. Una particolarità della metafase meiotica I è che i cinetocori
di ciascuna coppia di cromatidi fratelli sembrano essersi uniti. I microtubuli (4) del fuso centrale, che sono
ora collegati in modo preciso ai cinetocori di ciascuna coppia di cromatidi fratelli (5), puntano di conse-
guenza tutti nella stessa direzione. I chiasmi (6) sono ancora intatti. Esso ricoprono un ruolo importante
nel corretto allineamento dei cromosomi omologhi a livello equatoriale.
Il reticolo endoplasmatico (7) e l’apparato del Golgi (8) sono quasi completamente dissolti.
7. Anafase I
Nell’anafase I della meiosi, sono i cromosomi omologhi (1) a separarsi, invece dei cromatidi fratelli, come
avviene invece nella mitosi. Durante il processo si dissolvono i chiasmi, che fino a questo momento aveva-
no tenuto uniti i cromosomi materni e paterni.
Alcuni organismi mutanti, dove il crossing-over meiotico avviene solo entro certi limiti, presentano coppie
di cromosomi senza chiasmi. Di solito, queste coppie non sono completamente separate (non-disgiunzione)
e le cellule figlie risultanti hanno un cromosoma in meno o in più. Tali malformazioni vengono definite
aberrazioni cromosomiche numeriche e causano deformazioni.
La disgiunzione inizia in corrispondenza dei cinetocori (2), punto dove sono collegate le fibre del fuso cen-
trale. Da qui, i cromosomi vengono trascinati lentamente verso i centrioli (4), collocati ai poli della cellula,
spostandosi lungo i microtubuli (3) che creano un effetto di trazione man mano che si accorciano. I micro-
tubuli (5) non collegati ai cromosomi si allungano, aumentando così la distanza tra i centrioli ed estenden-
do la cellula. A livello equatoriale, diventa visibile lo stadio iniziale di un solco di clivaggio (6).
Il processo di crossing-over durante la profase e la distribuzione casuale dei cromosomi materni e paterni
ai poli della cellula portano a una diversificazione delle informazioni genetiche (vedere l‘introduzione).
8. Telofase I, citocinesi I, intercinesi, profase II e metafase II
Telofase I e citocinesi
Nella telofase I, il fuso si disintegra e a livello equatoriale si sviluppa una strozzatura (1). Viene inoltre a
crearsi una sottile membrana nucleare (2). Nel corso della successive fase di citocinesi, il corpo della cellula
si divide esattamente al centro, in corrispondenza della strozzatura tra i nuclei delle due nuove cellule
figlie (3). I nuclei figli contengono ciascuno l’assetto cromosomico materno e paterno leggermente alterato
attraverso il processo di crossing-over, dove il DNA è già presente in forma duplicata, ossia un cromosoma è
costituito da due cromatidi fratelli (4).
Il reticolo endoplasmatico (5) e l’apparato del Golgi (6) hanno riassunto la loro forma e le loro dimensioni
iniziali.
Alla fine della citocinesi, la prima divisione meiotica è completa.
35

®
ItalianoMeiosi
Intercinesi
La prima e la seconda divisione meiotica sono separate da un breve periodo di riposo (interfase). Tuttavia,
non avviene alcuna duplicazione dei cromosomi costituiti da due cromatidi (non è presente alcuna fase S).
Entrambi i cromatidi fratelli di ciascun cromosoma restano collegati dai centromeri (7).
Divisione meiotica II
La seconda divisione meiotica avviene esattamente come la mitosi (normale divisione nucleare e cellulare).
Viene perciò anche definita divisione equazionale. Poiché i cromosomi non sono stati nuovamente dupli-
cati durante la precedente intercinesi, la seconda divisione meiotica che ora segue include la riduzione
delle informazioni genetiche al solo assetto cromosomico aploide.
Profase II
La profase II è molto simile a quella della mitosi e avviene molto rapidamente in tutti gli organismi. La
permeabilità della superficie cellulare aumenta al fine di consentire l’assorbimento di liquido dall’esterno.
Il complesso microtubulare del citoscheletro si riorganizza, mentre la membrana nucleare si dissolve e il
fuso viene creato riordinando i microtubuli.
Metafase II
Nella metaphase II, I cromosomi vengono nuovamente posizionati a livello equatoriale e le due estremità
del fuso collocate ai due poli opposti della cellula, come nella metafase I. Una differenza sostanziale
rispetto a quest’ultima è che i due cinetocori si sono sviluppati in corrispondenza dei cromatidi fratelli, i
quali, in questo caso, puntano verso direzioni polari opposte.
9. Anafase II
Durante l’anafase II, i due cromatidi fratelli (1) di ciascun cromosoma vengono disgiunti, proprio come
nella mitosi.
La separazione inizia in corrispondenza dei cinetocori (2), punto dove sono collegate le fibre del fuso cen-
trale. Da qui, i cromosomi vengono trascinati lentamente verso i centrioli (4), collocati ai poli della cellula,
spostandosi lungo i microtubuli (3) che creano un effetto di trazione man mano che si accorciano. I micro-
tubuli (5) non collegati ai cromatidi si allungano, aumentando così la distanza tra i centrioli ed estendendo
la cellula. A livello equatoriale, diventa visibile lo stadio iniziale di un solco di clivaggio (6).
10. Telofase II e citocinesi II
Il clivaggio e la divisione delle due cellule prodotte durante la prima fase meiotica portano ora alla produ-
zione di quattro cellule aploidi (1) con diverse combinazioni genetiche derivanti da una distribuzione cro-
mosomica casuale e dal crossing-over. Ciò spiega perché i fratelli non sono identici: un bambino può avere
più tratti del padre, l’altro della madre. È inoltre possibile che riappaiano caratteristiche degli antenati.
36

®
37

®
38

®
39

®
40
