3B Scientific Animal cell model: инструкция
Раздел: Товары для здоровья
Тип:
Инструкция к 3B Scientific Animal cell model

R04
(1000523)

2

®
The Animal CellEnglish
Introduction
Cells in animal multicellular organisms principally only occur in groups of similar cells or together with
other differentiated cells, or embedded in the intercellular substrate (intercellular substance, extracellular
matrix). The surrounding environment of the unicellular and primitive multicellular organisms (the “pri-
mordial soup“, so to speak) also surrounds the cells of more complex highly organized animal (human)
organisms, and ensures its nutrition via the blood vessels that penetrate throughout the tissues (down to
the capillaries).
The following basic characteristics distinguish the cells of living organisms: they possess higher complexity
of organization than their surroundings, they can react to stimuli from within and from their environment,
and they have the ability to reproduce (reduplication).
Overview of the construction and function of cells
The cell membrane (plasma membrane) encloses the cell and also provides a barrier to the external envi-
ronment allowing the maintenance of its own internal environment. Within the cell, certain structures and
small organs (organelles, see list below) are also enclosed by a plasma membrane. The plasma membrane
itself consists of polar lipids that form a semi-permeable membrane. Thus the individual compartments
and organelles are separated from one another and from the specific molecules and ions they contain.
The plasma membrane is also connected to a fine framework of structural proteins, the filaments of the
cell skeleton (cytoskeleton). This cytoskeleton consists of fine actin filaments (7 nm diameter), hollow
microtubules (25 nm diameter) and, lying in between in diameter, the intermediary filaments. The micro-
tubules develop from an organization centre, usually the centriole. They are also responsible for transport
processes along their length, to and from the organization centre (directional active transport, which also
occurs in the axons of nerve cells). The centriole itself is an organelle consisting of two groups of tubes
perpendicular to one another, from which the microtubules extend – this also occurs in newly formed cells.
During cell division the separation of the chromosomes is carried out by the “marionette threads”, the
microtubules emanating from the centriole.
As the name implies, the cytoskeleton ensures overall stability for the cell along with a corresponding
degree of flexibility. Furthermore the cytoskeleton enables extreme versatility in the active movements of
the cell: from stretching out foot-like appendages (e.g. filopodia) to make major changes in shape of the
entire cell (also the basis of active muscle contraction for example) to active movement of the cell (cell
migration). Moreover, the elements of the cytoskeleton propagate the tension lines within a cell via the
so-called cell-cell connections (e.g. desmosomes, see below) to the neighbouring cells and so mechanically
connect different areas of cells e.g. in the epidermis of the skin – particularly clear in the prickle cells.
Within the cell-cell connections (intercellular contact) structures with predominantly mechanical function
(contact adhesion: zonula; punctum; fascia adhaerens; macula adhaerens = desmosome) can be distinguis-
hed from those with an active metabolic and electro-coupling function (nexus, macula communicans = gap
junction; synapse). Finally, there are cell connections that seal off the intercellular area (contact barrier:
zonula occludens). Connections to the extracellular membrane form focal contacts and to the basal memb-
rane the hemidesmosome.
All proteins, which make up the components of the cytoskeleton, are made by the “sewing machine“ of
the proteins, the ribosomes. These can be suspended in the cytoplasm or may be bound onto the vacuole
system of the rough endoplasmic reticulum (rough ER). Information is carried to the ribosomes from the
cell nucleus, where genetic information is stored on the chromosomes by means of the mRNA. The ribo-
some couples amino acid to amino acid to order and “sews” them onto a peptide or protein. Peptides and
proteins are further modified by auxiliary proteins within the ER, e.g. sugar groups may be added to the
protein (glycosylation). The smooth ER can synthesize lipids (cholesterol, triglycerides, steroid hormones),
lipoproteins and phospholipids. Furthermore the smooth ER makes fat-soluble compounds water-soluble
3

®
The Animal Cell
English
and thus detoxifies them. In certain types of cell (e.g. muscle cells) the smooth ER also serves as a store for
calcium ions.
The Golgi apparatus (GA) consists of groups of flat sac-like membranes (sacculi) und numerous small vesi-
cles. Here proteins newly synthesized in the ER are further modified. Specific sugar groups are synthesized
and coupled onto proteins, and also some proteins are cleaved. In addition polysaccharides with appropri-
ate sulfatization are synthesized and glycolipids are synthesized and joined together. These mature proteins
are packed into vesicles in the GA and made ready for transport. Thus the GA plays an important role in the
sorting and delivery of proteins (“Post Office of the cell“).
The GA is also connected with exocytotic events (release of vesicles) and fagocytosis (protrusion of the
plasma membrane with subsequent “pinching off” of the membrane together with the components that it
surrounds). Vesicles can also be actively enveloped and captured (endocytosis) and also receptors acting as
mediators can be brought in (pinocytosis). Membrane protrusions such as the microvilli (small finger-like
protrusions with little mobility), stereocilia (larger protrusions with relatively little mobility) and kinocilia
(flagellum-shaped protrusions for active cilia movement) will only be discussed in the course reading.
Vacuoles can be formed both endocytotically and by cell components that digest the contents of vesicles
(lysosomes).
Alongside the lysosomes in a cell are small organelles (peroxisomes), which principally oxidize fatty acids,
amino acids and uric acid and can detoxify potentially cytotoxic organic compounds.
Mitochondria are bacteria that entered unicellular organisms in primeval times and developed into so-
called symbionts. They possess their own genetic material and also ribosomes for the synthesis of their own
proteins. They have however become so intertwined with the genome of the host cell that they can no lon-
ger live independently. The benefit of this symbiosis is that the mitochondria can utilize oxygen (a principle
toxic substance for living cells, that first arose in the primeval atmosphere from the evolution of plants).
Thus carbohydrates, fatty acids and amino acids are oxidized to CO
and H
O by consumption of elemental
2
2
oxygen. In this way ATP, the universal energy carrier, is obtained for the cell. The mitochondrion possesses
a double membrane wall. ATP synthesis and the processes of the respiratory chain take place on the inter-
nal wall itself. Fatty acid oxidation and the citric acid cycle take place within the mitochondria matrix. Thus
the mitochondria can be described as the powerhouse of the cell.
The cell nucleus is the information centre for the cell. The information itself is distributed over 46 deoxy-
ribonucleic acid molecules (DNA). They are accommodated in the cell nucleus together with the histones
(capping proteins). The nucleus is typically more densely packed than the cytoplasm and is surrounded by
a nuclear double membrane (cistern of the ER) with defined channels (nuclear pores). The information for
protein synthesis is taken to the ribosomes by the messenger ribonucleic acid (mRNA), which is synthesized
on the gene segments of the DNA. This process is called transcription and produces copies of the DNA. The
ribosomal RNA is synthesized on specialized segments in the nucleoli (aggregations in the cell nucleus). In
addition there is close communication between the cytoplasm and the membrane receptors, so the cell
nucleus represents the central information and control unit of the cell.
Author: Prof. Dr. R.H.W. Funk, Institute of Anatomy, Dresden University of Technology
4

®
English The Animal Cell
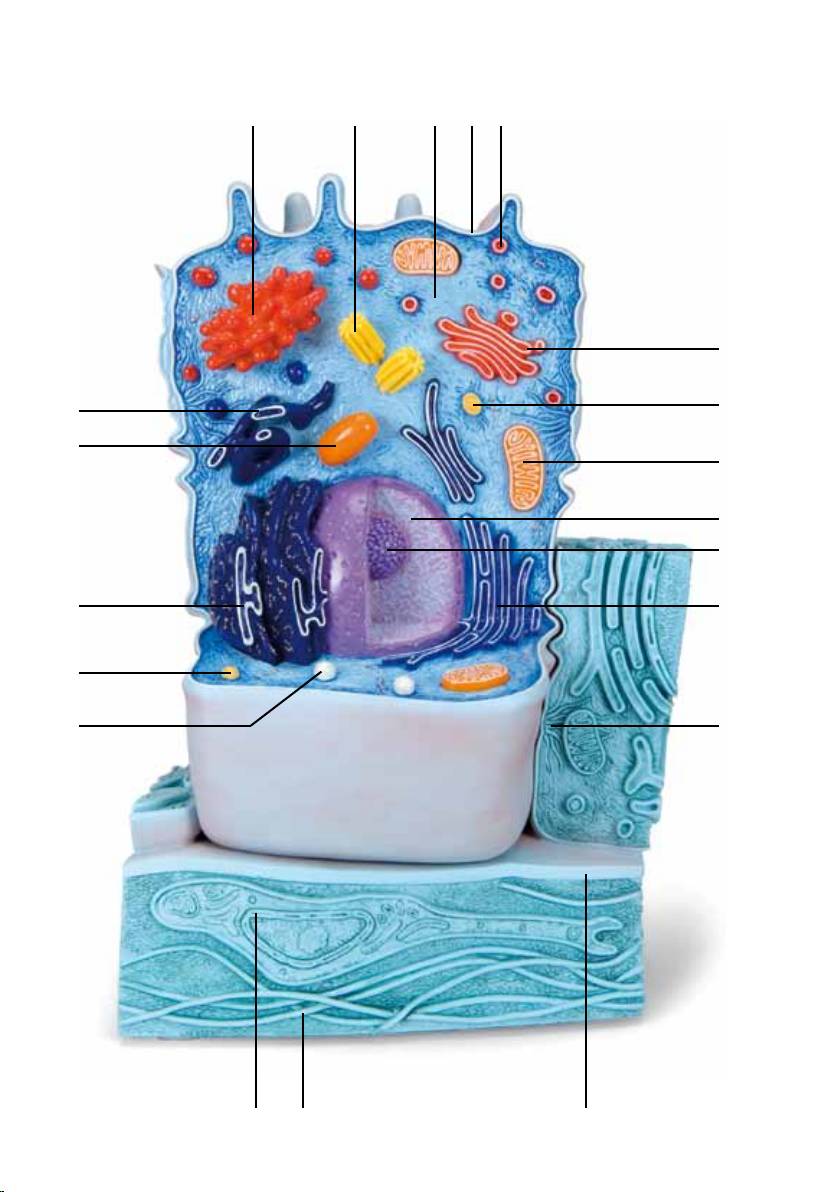
1 Cell nucleus
2 Nucleolus
3 Mitochondrion
4 Smooth endoplasmic reticulum (ER)
5 Desmosome (Macula adhaerens)
6 Basal membrane
7 Hemidesmosome
8 Collagen fibres
9 Fibroblast
10 Peroxisome
11 Lysosome
12 Rough endoplasmic reticulum (ER)
13 Mitochondrion
14 Smooth endoplasmic reticulum (ER)
15 Golgi Apparatus
16 Centriole
17 Cytosol with embedded filament of the cytoskeleton
18 Microvilli
19 Secretion vesicle
20 Golgi Apparatus
21 Lysosome
22 Zonula occludens
23 Zonula adhaerens
24 Desmosome (Macula adhaerens)
25 Microplica
5

®
DeutschDie tierische Zelle
Einleitung
Im tierischen mehrzelligen Organismus kommen Zellen grundsätzlich nur im Verbund von vielen gleich-
artigen bzw. unterschiedlich differenzierten Zellen, bzw. eingebettet in der zwischenzelligen Substanz
(Interzellularsubstanz, extrazelluläre Matrix) vor. Das Umgebungsmilieu der Einzeller und primitiven
Mehrzeller (gleichsam das „Urmeer“) umgibt so auch die Zellen in komplexen tierischen (menschlichen)
Organismen, wobei über die allseits das Gewebe durchdringenden Blutgefäße (bis herunter zu den
Haargefäßen, Kapillaren) die Ernährung gewährleistet wird.
Folgende Grundcharakteristiken zeichnen Zellen lebendiger Organismen aus: Sie sind höher komplex orga-
nisiert als ihre Umgebung, sie können auf Reize aus ihrem Inneren und ihrer Umgebung reagieren, schließ-
lich haben sie die Fähigkeit sich zu vermehren (Reduplikation).
Übersicht über Bau und Funktion der Zelle
Die Umhüllung der Zelle und auch die Barriere zur Umwelt, zur Aufrechterhaltung eines eigenen inne-
ren Milieus, wird durch die Zellmembran (Plasmamembran) erreicht. Auch innerhalb der Zelle werden
bestimmte Strukturen und kleine Organe (Organellen, siehe Aufzählung unten) durch die Plasmamembran
umhüllt. Die Plasmamembran selbst besteht aus polaren Lipiden, die eine semipermeable Membran
bilden. Daher können sich die einzelnen Kompartimente und Organellen voneinander bezüglich des
Gehalts bestimmter Moleküle und Ionen abgrenzen. Die Plasmamembran nimmt auch Verbindungen
mit einem feinen Gerüst von Strukturproteinen, den Filamenten des Zellskeletts (Zytoskelett) auf. Hier
findet man feine Aktinfäden (7 nm Durchmesser), röhrenförmige Mikrotubuli (25 nm Durchmesser) und
die im Durchmesser dazwischen (intermediär) gelegenen Intermediärfilamente. Die Mikrotubuli wach-
sen von einem Organisationszentrum, meist dem Zentriol aus. Dabei sind sie auch verantwortlich für
Transportvorgänge an ihnen entlang, zu und vom Organisationszentrum weg (gerichteter aktiver Transport
– auch in Axonen von Nervenzellen). Das Zentriol selbst ist ein Organell aus zwei senkrecht aufeinander
stehenden Gruppen von Röhren, von denen aus die Mikrotubuli ausstrahlen – auch bei neu entstehenden
Zellen. Bei der Zellteilung wird die Trennung der Chromosomen von den „Marionettenfäden“, der von den
Zentriolen ausgehenden Mikrotubuli, ausgeführt.
Wie der Name „Zytoskelett“ schon sagt, erfüllt dieses die Aufgaben, der Zelle insgesamt Stabilität und eine
entsprechende Flexibilität zu gewährleisten. Darüber hinaus ermöglicht das Zytoskelett die vielfältigsten
aktiven Bewegungen der Zelle: Vom Ausstrecken von füßchenförmigen Fortsätzen (z. B. Filopodien) über
starke Formveränderungen der gesamten Zelle (Grundlage z. B. auch der aktiven Muskelverkürzung) bis
hin zur aktiven Wanderung (Migration) der Zelle. Des Weiteren führen die Elemente des Zytoskeletts die
Spannungslinien innerhalb einer Zelle über die so genannten Zell-Zellverbindungen (z. B. Desmosomen,
s. u.) weiter an die Nachbarzellen und bilden so einen mechanischen Verbund von Zellarealen (z. B. in der
Epidermis der Haut – besonders deutlich bei den Stachelzellen).
Innerhalb der Zell-Zellverbindungen (Interzellulärkontakte) lassen sich funktionell solche, mit über-
wiegend mechanischer Funktion (Adhäsionskontakte: Zonula; Punctum; Fascia adhaerens; Macula
adhaerens = Desmosom) von solchen mit stoffwechselaktiver (metabolischer) und elektrisch-koppelnder
Funktion (Nexus, Macula communicans = Gap junction; Synapsen) unterscheiden. Schließlich existie-
ren noch Zellverbindungen, die den Interzellularraum abdichten (Barrierenkontake: Zonula occlu-
dens). Verbindungen zur extrazellulären Matrix bilden fokale Kontakte bzw. zur Basalmembran das
Hemidesmosom.
Alle Proteine, die die Bestandteile des Zytoskeletts ausmachen, werden von den „Nähmaschinen“ der
Proteine, den Ribosomen, geliefert. Diese können frei im Zytoplasma liegen oder an das Vakuolensystem
des rauhen Endoplasmatischen Retikulums (rauhes ER) gebunden sein. Mit der Information, die aus
dem Zellkern (dieser speichert die Erbinformation über die Chromosomen) über die mRNA übermittelt
wird, koppelt das Ribosom nun „nach Anleitung“ Aminosäure an Aminosäure und „näht“ diese zu einem
6

®
Die tierische ZelleDeutsch
Peptid bzw. Protein zusammen. Im Kompartiment des ER werden Peptide und Proteine weiter durch
Helferproteine modifiziert, so dass z. B. Zuckergruppen auf das Protein übertragen werden können
(Glykosylierung). Das glatte ER kann Lipide (Cholesterol, Triglyceride, Steroidhormone) und Lipoproteine
sowie Phospholipide synthetisieren. Darüber hinaus macht das glatte ER lipidlösliche Verbindungen was-
serlöslich und entgiftet sie dadurch. In bestimmten Zellarten (z. B. Muskelzellen) dient dem glatten ER auch
als Speicher für Kalziumionen.
Der Golgi-Apparat (GA) besteht aus Gruppen flacher sackförmiger Membranen (Sacculi) und zahlreichen
kleinen Vesikeln. In diesem System werden die im ER synthetisierten noch unreifen Proteine weiter ver-
ändert. Es werden bestimmte Zuckergruppen synthetisiert und an Proteine angekoppelt, auch manche
Proteine gespalten. Des Weiteren werden Polysaccharide mit entsprechender Sulfatierung synthetisiert
sowie Glycolipide synthetisiert und angeknüpft. Die reifen Proteine werden im GA in Vesikel verpackt
und transportfähig gemacht. So spielt der GA eine wichtige Rolle bei der Sortierung und Auslieferung von
Proteinen („Postamt der Zelle“).
Hier steht der GA auch mit den Prozessen der Exozytose (Ausschleusung von Vesikeln) und Apozytose
(Ausbuchtungen der Plasmamembran, die mit den von ihnen umschlossenen Komponenten abgeschnürt
werden) in Verbindung. Darüber hinaus können Vesikel aktiv eingeschnürt werden (Endozytose) und auch
Rezeptor vermittelt eingebracht werden (Pinozytose). Nur kursorisch soll auf die Membranausstülpungen
wie Mikrovilli (kleine fingerförmige Ausstülpungen mit geringer Beweglichkeit), Stereozilien (größere
Ausstülpungen mit relativ geringer Beweglichkeit), Kinozilien (geißelförmige Ausstülpungen für aktiven
Flimmerschlag) eingegangen werden.
Sowohl endozytotisch als auch über Zellbestandteile selbst können Vakuolen gebildet werden, die die
abgeschnürten Inhalte verdauen (Lysosomen).
Neben den Lysosomen liegen in einer Zelle kleine Organellen (Peroxisomen), die vor allem Fettsäuren,
Aminosäuren und Harnsäure oxidieren und potentiell zelltoxische organische Verbindungen entgiften kön-
nen.
Mitochondrien sind vor Urzeiten in Einzeller eingewanderte Bakterien, die sich zu so genannten
Symbionten entwickelten. Sie besitzen eine eigene Erbinformation und auch Ribosomen für die Synthese
von eigenen Proteinen. Sie haben sich jedoch so intensiv mit dem Genom der Wirtszellen verfloch-
ten, dass sie nicht mehr eigenständig lebensfähig sind. Der Nutzen dieser Symbiose besteht darin,
dass Mitochondrien Sauerstoff (eine für die lebenden Zellen prinzipiell giftige Substanz, die erst in der
Uratmosphäre durch die Evolution der Pflanzen entstanden ist) verbrennen können, d. h. Kohlenhydrate,
Fettsäuren und Aminosäuren werden unter Verbrauch von elementarem Sauerstoff zu CO
und H
O
2
2
oxidiert. Dadurch wird der universelle Energieträger ATP für die Zelle gewonnen. Das Mitochondrium
besitzt eine doppelte Membranwand. An der inneren Membran selbst ist die Atmungskette und die ATP-
Synthesekette untergebracht. Im Inneren der Mitochondrienmatrix laufen die Fettsäureoxidation und der
Zitratzyklus ab. Damit stellen die Mitochondrien die „Kraftwerke“ der Zelle dar.
Der Zellkern ist das Informationszentrum für die Zelle. Die Information selbst ist auf 46
Desoxyribonukleinsäuremolekülen (DNA) verteilt. Sie sind zusammen mit den Histonen (Halteproteinen)
im Zellkern untergebracht. Der Kern, der insgesamt dichter gepackt ist als das Zytoplasma, wird von
einer Kerndoppelmembran (Zisterne des ER) mit definierten Kanälen (Kernporen) umgeben. Durch die
Boten-Ribonukleinsäure (mRNA), die an den Genabschnitten der DNA systhetisiert wird (Transkription)
und dann Kopien der DNA enthält, wird die Information zur Proteinsynthese zu den Ribosomen gebracht.
In den Nucleoli (Verdichtungen im Zellkern) werden an speziellen Abschnitten die ribosomale RNA
(rRNA) synthetisiert. Darüber hinaus besteht eine enge Kommunikation mit dem Zytoplasma und den
Membranrezeptoren, so dass der Zellkern die zentrale Informations- und Steuereinheit der Zelle darstellt.
Autor: Prof. Dr. R.H.W. Funk, Institut für Anatomie T.U. Dresden
7

®
DeutschDie tierische Zelle
1 Zellkern (Nucleus)
2 Nucleolus
3 Mitochondrium
4 Glattes Endoplasmatisches Retikulum (ER)
5 Desmosom (Macula adhaerens)
6 Basalmembran
7 Hemidesmosom
8 Kollagene Fasern
9 Fibroblast
10 Peroxisom
11 Lysosom
12 Rauhes Endoplasmatisches Retikulum (ER)
13 Mitochondrium
14 Glattes Endoplasmatisches Retikulum (ER)
15 Golgi Apparat
16 Zentriol
17 Zytosol mit eingebetteten Filamenten des Zytoskeletts
18 Mikrovilli
19 Sekretvesikel
20 Golgi Apparat
21 Lysosom
22 Zonula occludens
23 Zonula adhaerens
24 Desmosom (Macula adhaerens)
25 Mikroplica
8

®
La Célula AnimalEspañol
Introducción
En los organismos pluricelulares, básicamente, las células se encuentran sólo en conjunto con muchas
otras células del mismo tipo, o de un tipo diferenciado, o incrustadas en la sustancia intercelular (matriz
extracelular). El medio que rodea a los unicelulares y los pluricelulares primitivos (algo así como un mar
originario) rodea también las células de organismos animales complejos (humanos), en donde, sobre todo,
los vasos sanguíneos que atraviesan el tejido (hasta llegar a los capilares) garantizan la nutrición.
Las células de los organismos vivos presentan las siguientes características básicas: Están organizadas de
forma más elevada y compleja que su entorno, pueden reaccionar a sus propios estímulos internos y a los
de su entorno. Por último, tienen la capacidad de reproducirse (reduplicación).
Nociones generales sobre la estructura y la función de la célula
La membrana celular (membrana plasmática) le proporciona una cubierta a la célula y forma una barrera
con su entorno, que le permite mantener aislado su propio medio interno. En el interior de la célula tam-
bién hay determinadas estructuras y órganos pequeños (organelos, mirar la enumeración abajo) que se
encuentran recubiertos por la membrana plasmática. La membrana plasmática está compuesta por lípidos
polares que forman una membrana semipermeable. Debido a esto, los compartimentos individuales y los
organelos pueden delimitar entre sí mismos el contenido de determinadas moléculas e iones. La membra-
na plasmática también tiene contactos con un fino andamio de proteínas estructurales y filamentos del
citoesqueleto. Aquí se encuentran los finos hilos de actina (7 nm de diámetro), los microtúbulos, llamados
así por su forma (25 nm de diámetro), y los filamentos intermedios que se encuentran en diámetros inter-
medios. Los microtúbulos crecen a partir de un centro de organización que, por lo general, es el centriolo.
También son responsables de los procesos de transporte que, a través de ellos, salen desde el centro de
organización o se dirigen hacia él (transporte activo y dirigido, también presente en los axones de las célu-
las nerviosas).
El centriolo, en sí mismo, es un organelo que se compone de dos grupos de tubos ubicados verticalmente
uno frente al otro, de los cuales se desprenden los microtúbulos, lo cual ocurre también en las células
recién formadas. Durante la división celular, los cromosomas se separan de los “hilos de marioneta” de los
microtúbulos que provienen del centriolo.
Como el nombre lo indica, el citoesqueleto tiene la tarea de garantizarle a la célula una estabilidad general
y una flexibilidad adecuada. Por otra parte, el citoesqueleto posibilita la multiplicidad de los movimientos
activos de la célula: desde la extensión de algunos apéndices con forma de pies pequeños (por ejemplo,
los filopodios), fuertes cambios en la forma de toda la célula (lo que, por ejemplo, constituye la base de la
contracción muscular activa) hasta un desplazamiento activo de la célula misma (migración). Además, los
elementos del citoesqueleto dirigen las líneas de tensión interna de la célula hacia las células vecinas por
medio de las llamadas uniones celulares (por ejemplo, los desmosomas, ver abajo) y forman una unión
mecánica de áreas de células (por ejemplo, en la epidermis de la piel, especialmente en las células del
estrato espinoso).
Dentro de las uniones de las células (contactos intercelulares) se pueden diferenciar funcionalmente aqué-
llas que poseen una función mecánica (contactos de adhesión: zónula, punctum, fascia adherente, mácula
adherente = desmosoma) de aquéllas que tienen un metabolismo activo (metabólicos) al igual que de
aquéllas con una función de acoplamiento eléctrico (nexos, mácula comunicante = uniones comunicantes,
sinapsis). Por último, también existen uniones celulares que hermetizan el espacio intercelular (contactos
de barrera: Zonula occludens). Las uniones con la matriz extracelular forman contactos focales como el
hemidesmosoma con la membrana basal.
Todas las proteínas que forman parte de los componentes del citoesqueleto son suministradas por los
“fabricantes” de proteínas: los ribosomas. Estos pueden encontrarse libremente en el citoplasma o unidos
al sistema de vacuolas del retículo endoplasmático granular. Los ribosomas usan la información que pro-
viene del núcleo celular (éste almacena la información hereditaria por medio de los cromosomas) y que es
transmitida a ellos por el mRNA (RNA mensajero) para acoplar “de acuerdo con las instrucciones” un ami-
noácido a otro aminoácido, hasta formar un péptido o una proteína. Las proteínas y péptidos son modifi-
9

®
EspañolLa Célula Animal
cados en los compartimentos del retículo endoplasmático por medio de proteínas auxiliares, de forma tal
que, por ejemplo, los grupos de azúcar puedan transmitirse a las proteínas (glicosilación). El retículo endo-
plasmático liso puede sintetizar lípidos (colesterol, triglicéridos, hormonas esteroides) y lipoproteínas como
los fosfolípidos. Además, trasforma en solubles en agua los compuestos solubles en lípidos, volviéndolos
no venenosas. En determinados tipos de células el retículo endoplasmático liso también puede almacenar
iones de calcio (por ejemplo, en las células musculares).
El aparato de Golgi se compone de grupos de membranas planas y en forma de saco (sacculi) y de muchas
vesículas pequeñas. En este sistema se modifican nuevamente las proteínas inmaduras que han sido sinte-
tizadas en el retículo endoplasmático. Aquí se crean determinados grupos de azúcar y se acoplan a las pro-
teínas; también se dividen algunas proteínas. Por otra parte, se sintetizan los polisacáridos con la sulfataci-
ón respectiva, y también los glicolípidos se sintetizan y acoplan. Las proteínas ya maduras son empacadas
en el aparato de Golgi volviéndose aptas para el transporte. El aparato de Golgi juega un papel importante
en la clasificación y suministro de proteínas (como si fuera una “oficina de correos de la célula”).
El aparato de Golgi está ligado a los procesos de exocitosis (exclusión de las vesículas) y fagocitosis (una pro-
minencia de la membrana plasmática rodea una sustancia y la introduce al interior de la célula). Las vesí-
culas pueden verse atrapadas activamente (endocitosis) y los receptores absorbidos también activamente
(pinocitosis).
Solo se abordarán de forma esquemática las extensiones de la membrana como las microvellocidades
(pequeñas prominencias con forma de dedo y movilidad mínima), estereocilios (grandes prominencias con
relativamente mínima movilidad), quinocilios (prominencias en forma de un cilio para una vibración activa
en forma de golpe).
Por medio de la endocitosis, o con partes de la célula misma, se pueden formar las vacuolas, que son
capaces de digerir el contenido introducido (lisosomas). Además de los lisosomas, hay en las células peque-
ños organelos (peroxisomas) que permiten la oxidación de ácidos grasos, aminoácidos y ácidos úricos, y
eliminan el veneno de las formaciones orgánicas potencialmente tóxicas para la célula.
Las mitocondrias son bacterias que, en tiempos inmemoriales, migraron al interior de los seres unicelu-
lares convirtiéndose en lo que conocemos como simbiontes. Poseen información hereditaria propia, al
igual que ribosomas para la síntesis de las proteínas también propias. Se mezclaron tan profundamente
con el genoma de su huésped que ya no son capaces de sobrevivir independientemente. La utilidad de esta
simbiosis se basa en el hecho de que las mitocondrias pueden quemar oxígeno (una sustancia, en princi-
pio, venenosa para la célula, que sólo surgió en la atmósfera primitiva gracias a la evolución de las plan-
tas), o sea, los carbohidratos, ácidos grasos y aminoácidos se oxidan en CO
y H
O por medio del oxígeno
2
2
elemental. Así se genera para la célula el ATP, el transportador universal de energía. La mitocondria posee
una membrana con pared doble. En la propia membrana interna se encuentra la cadena respiratoria y la
cadena sintetizadora de ATP. En el interior de la matriz de la mitocondria se produce la oxidación de los
ácidos grasos y el ciclo de los citratos. De esta manera, las mitocondrias pueden considerarse como el “tal-
ler de energía de la célula”.
El núcleo celular es el centro de información de la célula. La información se encuentra repartida en 46
moléculas de ácido desoxirribonucléico (ADN). Ellas se alojan junto con las histonas (proteínas de sostén)
en el núcleo celular. El núcleo es en general más denso que el citoplasma y está rodeado por una mem-
brana nuclear doble (cisterna del retículo endoplasmático) con canales definidos (poros nucleares). La
información sobre la síntesis de las proteínas se transmite a los ribosomas a través de los ácidos ribonucl-
éicos mensajeros (mRNA) que se sintetizan en algunos sectores del gen del ADN (transcripción) y que luego
contienen copias de ADN. Los ácidos ribonucléicos mensajeros (mARN) se sintetizan en algunos sectores del
gen del AND (transcripción) y contienen una copia del ADN, por medio de éste llega la información de la
síntesis de las proteínas a los ribosomas. En los nucleolos (masa del núcleo celular), en algunas zonas espe-
ciales, se sintetiza el ARN ribosomal (rARN). Además, existe una estrecha comunicación entre el citoplasma
10

®
Español La Célula Animal
y los receptores membranales, de tal forma que el núcleo representa el centro de información y dirección
de la célula.
Autor: Prof. Dr. R.H.W. Funk, Instituto de Anatomía de la U. T. de Dresden
1 Núcleo celular
2 Nucleolo
3 Mitocondria
4 Retículo endoplasmático liso (RE)
5 Desmosoma (mácula adherente)
6 Membrana basal
7 Hemidesmosoma
8 Fibras colágenas
9 Fibroblasto
10 Peroxisoma
11 Lisosoma
12 Retículo endoplasmático granular (RE)
13 Mitocondria
14 Retículo endoplasmático granular liso (RE)
15 Aparato de Golgi
16 Centriolo
17 Citosol con filamentos incrustados del citoesqueleto
18 Microvellocidades
19 Vesícula secretora
20 Aparato de Golgi
21 Lisosoma
22 Zónula ocludente
23 Zónula adherente
24 Desmosoma (mácula adherente)
25 Micropliegue
11

®
FrançaisLa cellule animale
Introduction
Dans l’organisme animal pluricellulaire, les cellules ne se présentent en principe que sous forme d’une
liaison de plusieurs cellules de même type ou sous forme de cellules à différenciation diverse. Les cellules
peuvent être également incluses dans la substance intercellulaire (substance intercellulaire, matrice extra-
cellulaire). Le milieu environnemental des organismes unicellulaires et des organismes pluricellulaires
primitifs (la « soupe primitive » pour ainsi dire) enveloppe également les cellules des organismes animaux
(humains) complexes. Leur alimentation est assurée par les vaisseaux sanguins se chargeant d’une irrigati-
on générale des tissus (jusqu’aux artères ciliaires, capillaires).
Les caractéristiques de base suivantes définissent les cellules d’organismes vivants : L’organisation de ces
cellules est plus complexe que celle de leur environnement. Elles peuvent réagir à des stimuli venant de
leur milieu interne ou de leur environnement, et elles disposent enfin de la faculté de se reproduire (répli-
cation).
Aperçu sur la structure et le fonctionnement de la cellule
La membrane cellulaire (membrane plasmique) forme l’enveloppe de la cellule et constitue également
une barrière contre l’environnement qui permet de préserver l’intégrité du milieu interne cellulaire. À
l’intérieur de la cellule elle-même, certaines structures et des petits organes intracellulaires (organelles, cf.
énumération ci-dessous) sont également enveloppés d’une membrane plasmique. La membrane plasmique
elle-même est composée de lipides polaires, formant une membrane semi-perméable. Les divers compar-
timents et organelles peuvent donc s’isoler les uns par rapport aux autres quant à la teneur de certains
ions et molécules. La membrane plasmique présente également des jonctions avec une ossature fine de
protéines structurales, les filaments du squelette cellulaire (cytosquelette). Nous y trouvons des filaments
fins d’actine (diamètre de 7 nm), des microtubules à structure tubulaire (diamètre de 25 nm) ainsi que les
filaments intermédiaires dont le diamètre présente une valeur intermédiaire. Les microtubules « poussent
» à partir d’un centre d’organisation, généralement depuis le centriole. Ils assument en outre les processus
de transport sur toute leur propre longueur, vers et depuis le centre d’organisation (transport directionnel
actif – également dans les axones de cellules nerveuses). Le centriole lui-même est un organelle composé
de deux groupements de tubes perpendiculaires les uns aux autres, à partir desquels les microtubules
rayonnent – également lors de la formation de nouvelles cellules. Lors de la division cellulaire, la sépa-
ration des chromosomes se fera au moyen des « ficelles de marionnette », les microtubules partant des
centrioles.
Comme son nom l’indique, les tâches assumées par le « cytosquelette » permettront une plus grande
stabilité générale de la cellule tout en conférant à cette dernière la flexibilité requise. Le cytosquelette
permet en outre les mouvements actifs les plus variés de la cellule : depuis l’extension des prolongements
pédiformes (tels que filopodes) en passant par des modifications importantes de l’ensemble de la forme
cellulaire (sur lesquelles se base par exemple le raccourcissement musculaire) jusqu’à la migration active
de la cellule. Par ailleurs, les éléments du cytosquelette transmettent les lignes de contrainte se présentant
à l’intérieur d’une cellule aux cellules voisines, via les jonctions cellule-cellule (telles que les desmosomes,
cf. ci-dessous), en formant une liaison mécanique, composée de zones cellulaires (telles que celles pré-
sentes dans l’épiderme de la peau – ce qui se manifeste visiblement dans le cas de la couche des cellules
épineuses).
À l’intérieur des jonctions cellule-cellule (contacts intercellulaires), il est possible de faire une distinction
fonctionnelle entre d’une part les jonctions assumant une fonction essentiellement mécanique (contacts
d’ancrage : zonula ; punctum ; fascia adhaerens ; macula adhaerens = desmosome) et entre d’autre
part les jonctions assumant une fonction métabolique active et de jonction électrique (nexus, macula
communicante = jonction communicante ; synapses). Enfin, il existe encore des jonctions cellulaires
assurant une étanchéification de l’espace intercellulaire (contacts au niveau de la barrière : zonula occlu-
12

®
Français La cellule animale
dens). Des contacts focaux forment des jonctions vers la matrice extracellulaire ou la membrane basale,
l’hémidesmosome.
Toutes les protéines, composant le cytosquelette, sont délivrées par les « machines à coudre » des proté-
ines, les ribosomes. Ces derniers peuvent circuler librement dans le cytoplasme ou être reliés au système
vacuolaire du réticulum endoplasmique rugueux (RE rugueux). Au moyen des informations transmises
depuis le noyau cellulaire (ce dernier stocke l’information génétique dans les chromosomes) via l’ARNm,
le ribosome couple alors « conformément au mode d’emploi » acide aminé à acide aminé et les « coud »
ensemble pour en faire un peptide ou une protéine. Dans le compartiment du RE, des protéines auxiliaires
se chargeront de poursuivre la modification des peptides et des protéines afin que des groupements sucrés
puissent être par exemple transférés à la protéine (glycosylation). Le RE lisse peut réaliser la synthèse de
lipides (cholestérol, triglycérides, hormones stéroïdiennes) et de lipoprotéines ainsi que celle de phospho-
lipides. Le RE lisse se chargera en outre de rendre composés liposolubles solubles dans l’eau, ce qui les
détoxiquera. Dans certains types de cellules (telles que cellules musculaires), le RE lisse joue également le
rôle d’un réservoir pour les ions calcium.
L’appareil de Golgi (AG) comprend des groupements de membranes plates en forme de sac (saccules) ainsi
que beaucoup de petites vacuoles. C’est au niveau de ce système que les protéines synthétisées dans le RE
et qui n’y sont pas encore arrivées à maturité subiront d’autres modifications. Certains groupements sucrés
seront synthétisés et couplés à des protéines, quelques protéines seront également scindées. Nous assistons
par ailleurs à une synthèse de polysaccharides avec une sulfatation correspondante ainsi qu’à une synthèse
et un attachement de glycolipides. Dans l’AG, les protéines arrivées à maturité seront emballées dans des
vacuoles et préparées au transport. L’AG joue donc un rôle important lors du tri et de la distribution des
protéines (« bureau de poste de la cellule »).
Dans ce cas, l’AG est également en rapport avec les processus de l’exocytose (rejet de vacuoles à l’extérieur)
et ceux de l’apocytose, le clivage de composants cellulaires (saillies de la membrane plasmique, étranglées
au moyen des composant les entourant). Des vacuoles peuvent en outre être activement étranglées (endo-
cytose) et des récepteurs intermédiaires également mis en place (pinocytose). Nous ne voulons traiter ici
que brièvement les excroissances de la membrane sous forme de microvillosité (petites excroissances à
mobilité restreinte), les stéréocils (excroissances de plus grande taille à mobilité relativement restreinte) et
les kinocils (excroissances de forme flagellée permettant une action sur le battement ciliaire).
Des vacuoles, digérant le matériel étranglé (lysosomes), pourront se former aussi bien par endocytose que
par les composants cellulaires eux-mêmes.
Une cellule contient, outre les lysosomes, de petites organelles (péroxisomes) pouvant avant tout oxyder
des acides gras, des acides aminés et des acides uriques et pouvant potentiellement détoxiquer des compo-
sés organiques toxiques au niveau cellulaire.
Les mitochondries sont des bactéries, ayant émigré, il y a des millions d’années, dans des organismes
unicellulaires et s’étant transformées en symbiontes. Ces mitochondries disposent d’une information géné-
tique propre et également de ribosomes leur permettant de synthétiser leurs propres protéines. Mais ces
bactéries se sont cependant alliées si étroitement au génome des cellules hôtes qu’elles ne sont plus auto-
nomement viables. Cette symbiose présente l’avantage que les mitochondries peuvent brûler de l’oxygène
(une substance en principe toxique pour les cellules vivantes, apparue seulement à la suite de l’évolution
végétales dans l’atmosphère primaire), en d’autres mots, des hydrates de carbone, des acides gras et des
acides aminés seront oxydés pour être transformés en CO
et H
O, en faisant appel à de l’oxygène élémen-
2
2
taire. Ce qui permettra de faire bénéficier la cellule de l’A.T.P, porteur universel d’énergie. La mitochondrie
présente une paroi membranaire double. La chaîne respiratoire permettant la synthèse de l’A.T.P se trouve
sur la membrane interne elle-même. L’oxydation des acides gras et le cycle de Krebs prennent place dans la
matrice des mitochondries. Les mitochondries représentent donc les « centrales électriques » de la cellule.
Le noyau cellulaire est le centre d’informations de la cellule. Les informations elles-mêmes sont réparties
13

®
FrançaisLa cellule animale
sur 46 molécules d’acide désoxyribonucléique (ADN). Qui se trouvent avec les histones (protéines d’ancrage)
dans le noyau cellulaire. Le noyau, dont la densité est dans l’ensemble plus compacte que celle du cyto-
plasme, est enveloppé d’une membrane nucléaire double (citerne du RE) présentant des canaux définis
(pores nucléaires). Les informations concernant la synthèse protéinique seront transmises aux ribosomes
par l’acide ribonucléique messager (ARNm), synthétisé sur les segments génétiques de l’ADN (transcription)
et contenant ensuite des copies de l’ADN. Dans les nucléoles (condensations dans le noyau cellulaire), la
synthèse de l’ARN ribosomique (ARNr) se fera au niveau de segments spéciaux. Il existe en outre une com-
munication étroite avec le cytoplasme et les récepteurs de la membrane, ce qui fait donc du noyau cellu-
laire l’unité centrale d’information et de commande de la cellule.
Auteur : Professeur Dr. R.H.W. Funk, Institut d’anatomie de l’Université Technique de Dresde
1 Noyau cellulaire (nucléus)
2 Nucléole
3 Mitochondrie
4 Réticulum endoplasmique lisse (RE)
5 Desmosome (Macula adhaerens)
6 Membrane basale
7 Hémidesmosome
8 Fibres collagènes
9 Fibroblaste
10 Péroxysome
11 Lysosome
12 Réticulum endoplasmique rugueux (RE)
13 Mitochondrie
14 Réticulum endoplasmique lisse (RE)
15 Appareil de golgi
16 Centriole
17 Cytosol avec filaments inclus du cytosquelette
18 Microvillosité
19 Vacuole de sécrétion
20 Appareil de golgi
21 Lysosome
22 Zonula occludens
23 Zonula adhaerens
24 Desmosome (Macula adhaerens)
25 Microplica
14

®
Português A célula animal
Introdução
Nos organismos animais pluricelulares as células geralmente sempre se encontram em associação com
muitas outras do mesmo tipo ou diferenciadas em diversas formas, integradas dentro de uma substância
entre as células (substância intercelular, matriz extracelular). O meio ambiente dos organismos unicelulares
e dos pluricelulares primitivos (parecido ao „mar originário“) envolve assim as células nos organismos ani-
mais complexos (humanos), sendo que a alimentação é garantida pelos vasos sangüíneos que atravessam o
tecido por todo lado (até os capilares).
As seguintes caraterísticas básicas definem os organismos vivos: eles tem uma organização mais complexa
do que o seu entorno, podem reagir a excitações internas e oriundas do seu meio ambiente externo, e
finalmente, eles têm a habilidade de se reproduzir (reduplicação).
Noções gerais sobre a constituição e o funcionamento da célula
O envoltório da célula, que é também uma barreira frente ao meio ambiente para poder criar um meio
interno próprio, é constituído pela membrana celular (a membrana plasmática). Também dentro da célula
existem algumas estruturas e pequenos órgãos (orgânulos, veja lista abaixo) que são envoltos de membra-
na plasmática. A própria membrana plasmática é feita de lipídios polares que formam uma membrana
semipermeável. Por isso, cada compartimento e orgânulo fica separado do outro conforme o conteúdo em
certas moléculas e íons. A membrana plasmática também tem contatos por meio de uma fina construção
de proteínas estruturais, os filamentos do esqueleto da célula (o citoesqueleto). Aqui encontram-se finos
filamentos de actina (7 nm de diâmetro), pequenos microtúbulos em forma de tubo (25 nm de diâmetro), e
os filamentos intermediários, intermediários pelo tamanho do diâmetro. Os microtúbulos crescem à partir
de um centro de organização, em geral o centríolo. Estes são responsáveis pelos transportes ao longo deles,
para outros centros de organização e desde eles (Transporte dirigido ativo – também presente nas axonas
das células nervosas). O centríolo em si é um orgânulo feito de dois grupos de tubos perpendiculares uns
aos outros, do qual partem os microtúbulos, também em células ainda em formação. Durante a divisão
celular, a divisão dos cromossomas é realizada através dos microtúbulos que saem dos centríolos.
Como o nome „citoesqueleto“ já indica, ele cumpre a função de garantir a estabilidade geral da célula,
assim como a necessária flexibilidade. Além disso, o citoesqueleto possibilita os mais variados movimen-
tos ativos da célula: desde a projeção de pontas em forma de pezinhos (por ex., os filopódios), ou fortes
alterações na aparência geral de toda a célula (a base, por ex., do encurtamento ativo dos músculos), até o
deslocamento ativo (migração) da célula. Pelo mais, os elementos do citoesqueleto conformam as linhas de
tensão dentro da célula e a conectam com outras células através da chamada comunicação célula a célula
(por ex., desmossomas), formando assim uma associação mecânica de zonas celulares (por ex., na epider-
me cutânea – particularmente visível nas células espinhosas).
Dentro das conexões de célula a célula (contatos intercelulares) distingue-se funcionalmente entre as
que cumprem funções mecânicas (contatos por adesão: zônula; junção intermediária (Fascia adhaerens);
mácula de adesão = desmossoma) das que cumprem funções ativas ligadas a processos do metabolismo
(metabólicas) ou uma função de conexão elétrica (nexo, junção comunicante = gap junction; sinapses)
Finalmente, existem também conexões celulares que vedam o espaço intercelular (contatos de barreira:
zônula de oclusão). As conexões com a matriz extracelular são formadas por contatos focais, como o
hemidesmossoma com a membrana basal.
Todas as proteínas que formam os componentes do citoesqueleto são fornecidas pelas „máquinas de costu-
ra“ das proteínas, os ribossomas. Estes podem estar livres no citoplasma ou estar associados ao sistema de
vacúolos do retículo endoplasmático rugoso (RE rugoso). Com as informações contidas no núcleo da célula
(este armazena a informação hereditária por meio dos cromossomas) e transmitidas pelo mRNA, o ribosso-
ma junta aminoácido com aminoácido conforme „as instruções“ e os „costura“ para formar um peptídeo
ou uma proteína. No compartimento do RE, os peptídeos e as proteínas são modificados por proteínas
15

®
A célula animal Português
auxiliares de modo a que grupos de açúcares, por exemplo, possam ser transferidos para a proteína (gli-
curonidação). O RE liso pode sintetizar lipídios (colesterol, triglicerídeos, hormônios esteróides) e lipopro-
teínas, assim como fosfolipídios. Além disto, o RE liso torna associações solúveis em lipídios em solúveis em
água, desintoxicando-as assim. Em certos tipos de células (por ex., nas células musculares) o RE liso serve
de armazém para íons de cálcio.
O complexo de Golgi (CG) consiste num grupo de membranas em forma de sacos achatados (sáculos) e
numerosas pequenas vesículas. Neste sistema, as proteínas sintetizadas no RE e ainda não maduras são
transformadas. Grupos específicos de açúcares são sintetizados e associados a proteínas, algumas proteínas
também são divididas. A continuação, são sintetizados polissacáridos com a sulfatação correspondente,
assim como glicolipídios são sintetizados e acoplados. As proteínas maduras são embaladas no CG numa
vesícula e preparadas para o transporte. Assim, o CG cumpre um papel importante na seleção e distribu-
ição de proteínas („o correio da célula“).
Aqui, o CG também está relacionado com os processos da exocitose (expulsão de vesículas) e da fagocitose
(invaginação da membrana que é logo fechada como um saco junto com os componentes assim abrangi-
dos). Além disso, vesículas podem ser fechadas ativamente (endocitose) e receptores podem ser absorvidos
ativamente (pinocitose). Só se abordarão de forma esquemática as extensões da membrana como os micro-
vilos (pequenas extensões em forma de dedo com pouca mobilidade), estereocílios (extensões maiores
com mobilidade relativamente pequena), cílios (extensões maiores em forma de chicote para vibrar ativa-
mente).
Tanto por endocitose como também por componentes da própria célula, podem formar-se vacúolos que
digerem os conteúdos absorvidos (lisossomas).
Além dos lisossomas encontram-se numa célula pequenos orgânulos (os peroxissomas), os quais oxidam
principalmente ácidos grassos, aminoácidos e ácido úrico e assim podem decompor formações químicas
orgânicas tóxicas para a célula.
As mitocôndrias são bactérias que em tempos primordiais migraram para dentro de seres unicelulares
tornando-se ditos simbiontes. Elas possuem uma informação hereditária própria assim como ribossomas
para a síntese de proteínas próprias. Elas se mesclaram tão profundamente com o genoma do hóspede que
elas não são mais capazes de sobreviver independentemente. A utilidade desta simbiose consiste no fato
que as mitocôndrias podem queimar o oxigênio (uma substância, em princípio, venenosa para a célula
que só surgiu na atmosfera primitiva graças à evolução das plantas), ou seja, os carboidratos, ácidos gras-
sos e aminoácidos são oxidados em CO
e H
O utilizando oxigênio elementar. Assim é produzido para a
2
2
célula o transportador universal de energia ATP. A mitocôndria possui uma membrana com parede dupla.
Na própria membrana interna encontram-se a corrente respiratória e a corrente sintetizadora de ATP. No
interior da matriz da mitocôndria ocorrem a oxidação dos ácidos grassos e o ciclo dos citratos. Com isto, as
mitocôndrias são as „usinas de energia da célula“.
O núcleo celular é o centro de informações da célula. A informação em si encontra-se distribuída em 46
moléculas de ácido desoxiribonucléico (DNA). Elas se encontram no núcleo junto com os histonos (pro-
teínas de suporte). O núcleo é geralmente mais denso do que o citoplasma, ele é envolto de uma membra-
na nucléica dupla (cisterna do RE) com canais definidos (poros nucléicos). Através dos ácidos ribonucléicos
mensageiros (mRNA) que são sintetizados nos trechos genéticos do DNA (transcrição) e que logo contêm
cópias do ADN, a informação sobre a síntese das proteínas é levada aos ribossomas. Nos nucléolos (densi-
ficações no núcleo) é sintetizado o RNA ribossomático (rRNA) em trechos especiais. Além disso, existe uma
comunicação próxima entre o citoplasma e os receptores da membrana, de modo que o núcleo representa
o centro de informações e a unidade de comando da célula.
Autor: Prof. Dr. R.H.W. Funk, Instituto de Anatomia da T.U. Dresden
16

®
A célula animalPortuguês
1 Núcleo
2 Nucléolo
3 Mitocôndria
4 Retículo endoplasmático liso (RE)
5 Desmossoma (mácula de adesão)
6 Membrana basal
7 Hemidesmossoma
8 Fibras colágenas
9 Fibroblasto
10 Peroxissoma
11 Lisossoma
12 Retículo endoplasmático rugoso (RE)
13 Mitocôndria
14 Retículo endoplasmático liso (RE)
15 Aparelho de Golgi
16 Centríolo
17 Citosol com filamento do citoesqueleto integrados
18 Microvilos
19 Vesícula secretora
20 Aparelho de Golgi
21 Lisossoma
22 Zônula de oclusão
23 Zônula de adesão
24 Desmossoma (mácula de adesão)
25 Microprega
17
15 16 17 18 19
20
21
14
13
3
1
2
12
4
11
10
5
689
18

22
23
24
25
7
19
19
- 1
- 2
